Mannosylated Fluorescent-DiO Macrophage Depletion Kit
SKU# CLD-8916
Description
Mannose receptor targeting by mannosylated liposomes has been demonstrated for a variety of mannosylated lipid conjugates in a variety of liposome morphologies and compositions in several different in vitro and in vivo models. A very large number of publications is about using a hydrophobic derivative of mannose (4-aminophenyl alpha-D-mannopyranoside) rather than using a mannosylated lipid in clodronate liposomes. This is mainly due to the high cost and complexity of synthesizing and conjugating mannose to lipid. 4-aminophenyl alpha-D-mannopyranoside is commercially available and far less expensive than synthesizing mannose conjugated lipid.
Why mannose? Mannose is one of the carbohydrate components of many bacterial and viral cell surfaces; therefore, the ever-efficient, highly redundant immune system has evolved multiple mechanisms for identifying pathogens based on mannose recognition. The animal and plant kingdoms likewise utilize carbohydrate recognition signaling mechanisms including mannose residues. Many publications evaluate other carbohydrates as targeting mechanisms for various cell types, however mannose targeting to phagocytes appears to be one of the more specific mechanisms identified to date. Mammalian cell surface identification molecules based on mannose binding, such as the ICAM family of leukocyte adhesion molecules, target the SIGN family of mannose receptors to accomplish self-recognition in vivo.
A well-known and cited study by Umezawa & Eto [1] demonstrates that liposomes containing aminophenyl mannoside were most efficiently incorporated into the mouse brain across the blood brain barrier. The radiolabeled liposomes bearing aminophenyl-alpha-D-mannopyranoside were maximally incorporated into the mouse brain after 48 hours, whereas in the spleen and liver, these radioactivities were maximum after 12 hours. The studies also showed that liposomes were most incorporated was glial cells rather than neuronal cell. The subcellular fractionation study indicates that mannose labeled liposomes are incorporated into lysosomes rich fraction both in liver and brain.
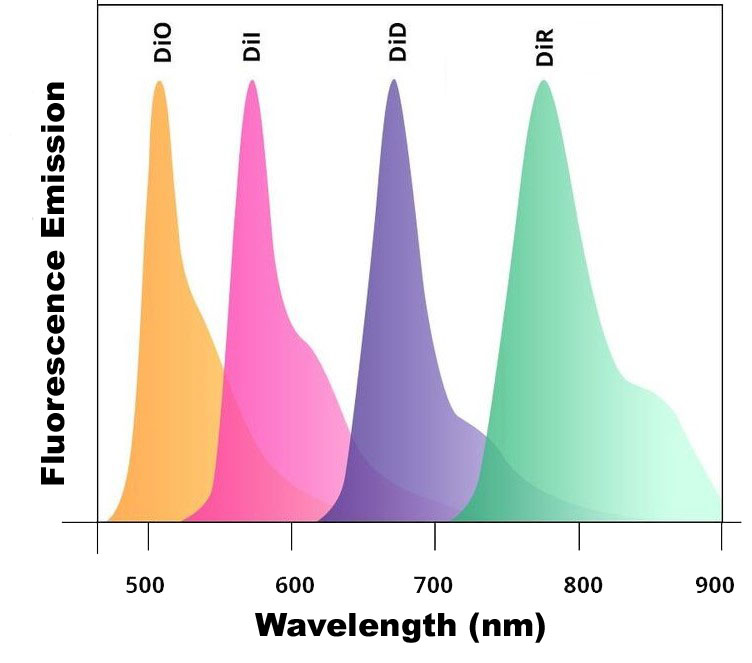
There are five mannosylated fluorescent control liposome products (m-Fluoroliposome®) for m-Clodrosome® (mannosylated clodronate liposomes). All five mannosylated fluorescent liposomes incorporate a lipophilic dye inside their membranes. They are insoluble in water; however, their fluorescence is easily detected when incorporated into membranes. DiI, DiO, DiD, DiR and DiA cover a wide range of excitation and emission wavelengths from 300s to 900s. DiI and DiO have fluorescence excitation and emission maxima separated by about 65 nm, facilitating two-color labeling. The emission spectrum of DiA is very broad, allowing it to be detected as green, orange, or even red fluorescence depending on the optical filter used. DiI, DiO, DiD and DiR belong to the dialkylcarbocyanines family of compounds. The spectral properties of the dialkylcarbocyanines are largely independent of the lengths of the alkyl chains but are instead determined by the heteroatoms in the terminal ring systems and the length of the connecting bridge. They have extremely high extinction coefficients, moderate fluorescence quantum yields, and short excited state lifetimes in lipid environments (~1 ns). The fluorescence spectrum of each dye is shown below.
You can choose the m-Fluoroliposome® based on the type of the fluorescent equipment and filters that you use in your lab. Mannosylated clodronate liposomes cannot be made fluorescent simply due to the potential for inaccurate and/or uninterpretable data being generated by labelled m-Clodrosome®. For more information, please refer to the technical note section.
Formulation Information
Clodrosome® Liposomal Clodronate Suspension
| Lipid Composition | Concentration (mg/ml) | Concentration (mM) | Molar Ratio Percentage |
|---|---|---|---|
| L-alpha-Phosphatidylcholine | 18.8 | 24.3 | 70 |
| Cholesterol | 4.2 | 10.9 | 30 |
| Total | 23 mg/ml | 35.1 mM | 100 |
| Encapsulated Drug | Concentration |
|---|---|
| Clodronate ((Dichloro-phosphono-methyl)phosphonate), Disodium Salt | 18.4* mM |
| * Depending on the type of the clodronate salt, its concentration (mg/ml) varies. If tetra hydrate salt is used, the concentration of the encapsulated drug will be ~7 mg/ml, and if a non-hydrated salt is used, the concentration will be ~5 mg/ml. | |
Fluoroliposome®-DiO
| Lipid Composition | Concentration (mg/ml) | Concentration (mM) | Molar Ratio Percentage |
|---|---|---|---|
| L-alpha-Phosphatidylcholine | 18.8 | 24.3 | 70 |
| Cholesterol | 4.2 | 10.9 | 30 |
| Total | 23 mg/ml | 35.1 mM | 100 |
| Mannosylation | Concentration |
|---|---|
| 4-Aminophenyl-alpha-D-mannopyranoside | 9.53 mol% |
| Fluorescent Dye | Excitation/Emission (nm) | Concentration (mg/ml) | Concentration (mM) |
|---|---|---|---|
3,3'-Dilinoleyloxacarbocyanine Perchlorate (DiO)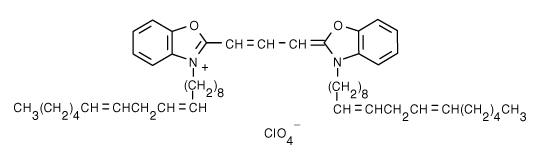 | 484/501 | 0.0625 | 0.071 |
| Buffer and Liposome Size | Specification |
|---|---|
| Buffer | Phosphate Buffered Saline |
| pH | 7.4 |
| Liposome Size | 1.5-2 µm |
Technical Notes
- To reach bloodstream-accessible, mannose-receptor positive cells outside the liver, a significant number of liposomes will have to escape first-pass uptake by the liver and spleen, so that the target cells are exposed to a higher concentration of mannosylated liposomes from the blood. One strategy that has been used to ensure that liposomes escape the liver and spleen is known as reticuloendothelial system (RES) blockade in which animals are pre-dosed with a sufficient quantity of liposomes to temporarily saturate the phagocytic cells of the blood, liver and spleen, also known as the reticuloendothelial system (RES) or the mononuclear phagocyte system (MPS). This sufficient quantity is dependent upon the liposome type and composition as well as the species being dosed; the pre-dosed liposomes do not necessarily need to be the same type or composition as the therapeutic or diagnostic liposomes avoiding the RES. Soon after this pre-dose is cleared from the bloodstream (usually within a couple of hours), the liposomes of interest are dosed. Since the RES is involved in digesting the previous dose of liposomes, the subsequently dosed liposomes will remain in the circulation much longer thus be much more likely to bind to their target site outside the RES including those phagocytic cells which are accessible, but are not usually exposed to a higher concentration of liposomes.
- While RES blockade is usually thought of as saturating phagocytic cells, it has been shown that opsonin-binding by liposomes is a saturable phenomenon. Therefore, part of RES blockade may involve serum depletion of complement and other opsonins known to coat liposomes. In the current application, removal or reduction in the concentration of soluble mannose-receptors may further increase the probability of a mannosylated liposome being able to interact with mannose receptors on the target cell. Therefore, if the goal is to deplete a target subset of mannose-receptor + cells which may not normally be exposed to a substantial number of mannosylated liposomes, pre-dosing with mannosylated clodronate liposomes, in order to both saturate the blood, liver and spleen phagocytes and reduce the concentration of opsonins including soluble mannose receptors, should increase the number of subsequently dosed mannosylated clodronate liposomes available to this target subset hypothetically resulting in increased uptake and depletion by these targeted cells.
- The issue with fluorescent Clodrosome® has to do with the potential for inaccurate and/or uninterpretable data being generated by labelled Clodrosome®. When Clodrosome® induces macrophage apoptosis, the fluorescent lipid incorporated into the Clodrosome® that is disrupted and metabolized in the phagolysosome will be dispersed among the residual apoptotic bodies which are subsequently phagocytosed by other macrophages. Therefore, fluorescent lipid may be detected in phagocytic cells which never phagocytosed Clodrosome® especially when FACS or fluoroscopy are utilized to detect fluorescent cells (FACS) or fluorescence levels in a tissue homogenate (fluoroscopy). Another potential artifact arises from fluorescent lipid remaining in the extracellular “garbage”, which has not yet been cleared by other phagocytes, generating a high background fluorescence. However, experienced confocal microscopist may be able to differentiate between the punctate fluorescence resulting from fluorescent intact liposomes versus the more diffuse fluorescence characteristic of disrupted liposomes and some have successfully used fluorescent clodronate liposomes to visualize the cellular location of these liposomes by confocal microscopy in vivo [2]. A further complicating factor is that published data varies widely as to exactly when clodronate liposomes begin to induce apoptosis in macrophages. Mönkönnnen et al. show that macrophage death is measurable within the first hour after clodronate liposome treatment on RAW264 cells in vitro [1], while others have reported no signs of macrophage apoptosis until several hours after treatment in vivo. The variability in the data is likely due to different liposomal formulations of clodronate as well as the vastly different experimental conditions. Therefore, as with most biological studies, especially those involving liposomes, the amount of time between treating the animal or cells with clodronate liposomes and the onset of apoptosis will need to be established in each experimental model. If the nature of the research demands that Clodrosome® be tracked rather than the control, Encapsula can provide DiI-labelled Clodrosome® upon request, and assuming that the Clodrosome® distribution can definitively be assessed prior to the onset of apoptosis, clear and valid data on the biodistribution of fluorescent Clodrosome® should be obtainable. Still, for most purposes, Fluoroliposome® (fluorescent control liposomes) will provide the required data with far fewer potential artifacts.
- When monitoring monocyte uptake in vivo in normal animals, the circulating monocytes may “disappear” or show reduced counts within the first 2 h post-injection due to margination of the monocytes post-liposome phagocytosis. These cells will re-enter the circulation within a few hours. Sunderkötter et al. demonstrate this phenomenon and discuss the behavior in detail. Also consider that circulating monocytes have a lifetime of about 24 h so labeled monocytes will be continually leaving the circulation, even in normal animals, due to aging of the monocytes [3].
- When animals or cells are treated with Clodrosome®, phagocytic cells recognize the liposomes as invading foreign particles and proceed to remove the liposomes from the local tissue or serum via phagocytosis. The liposomes then release clodronate into the cytosol resulting in cell death. Unencapsulated clodronate cannot cross the cell membrane to initiate cell death.
- Encapsome® control liposomes are recognized and phagocytosed by the same mechanism as Clodrosome®. Since the control liposomes do not contain clodronate, the phagocytic cells are not killed. However, phagocytes do respond to the ingestion of the control liposomes by cytokine secretion, temporary suspension of phagocytic activity and other responses described in the literature.
- The product must be removed from the vial using sterile technique. Do not use if sterility is compromised. This is particularly important if a single vial is accessed multiple times over several weeks. The product should not be used more than 60 days after receipt, even if unopened.
- Liposomes may settle when left undisturbed for more than a few hours. Immediately prior to use, in order to ensure a homogeneous liposome suspension, slowly invert the vial several times until the suspension appears homogeneous by visual inspection. Vigorous or erratic shaking will not damage the liposomes but may induce foaming and bubble formation making it more difficult to accurately measure the desired dosage.
- If the personnel performing intravenous injections are not experienced in or familiar with, precautions for injecting larger volumes (~10% animal weight in ml), viscous liquids or particulate suspensions, consider having extra animals available in case serious injection-related adverse events occur. Dose control animals first to become familiar with large volume injections.
- Within hours after systemic administration of Clodrosome®, animals begin to lose important components of their immune system. Standard animal handling and housing protocols are not suitable for immunocompromised animals. Even when such precautions are taken, monitor the general health of each animal for opportunistic infections unrelated to the experimental protocol. There is no inherent toxicity to the product at the recommended dose levels.
- When dosing intravenously, use standard precautions for dosing larger volumes to animals including the following: a) warm product to room temperature prior to dosing; b) ensure that all air bubbles are removed from the syringe prior to dosing. Intravenous injection of air bubbles may result in air emboli which can kill or seriously injure animals; c) inject product at a slow, steady rate of no more than 1 ml/min; d) decrease infusion rate if animals display any atypical reactions such as unusual agitation.
- Infusion-related adverse reactions usually involve the animal gasping for air or other seizure-like movements. Animals often recover with no apparent permanent injury, but any potential effects on experimental results must be assessed by the researcher.
- Liposomes should be kept at 4°C and NEVER be frozen.
Dosage
Appearance
m-Clodrosome® is a white milky suspension, and m-Fluoroliposome®-DiO is a yellow liquid suspension, both made of large micro size multilamellar liposomes. Due to their large size, some liposomes might settle to the bottom of the vial. If left sitting idle in the refrigerator, m-Fluoroliposome®-DiO will phase separate and form pellets in the bottom of the vial, leaving a clear solution on top. m-Clodrosome® might do the same only not as severely. Therefore, both should be shaken to form a homogeneous solution prior to use.
Educational Videos
Ordering/Shipping Information
- All liposome based formulations are shipped on blue ice at 4°C in insulated packages using overnight shipping or international express shipping.
- Liposomes should NEVER be frozen. Ice crystals that form in the lipid membrane can rupture the membrane, change the size of the liposomes and cause the encapsulated drug to leak out. Liposomes in liquid form should always be kept in the refrigerator.
- Clients who order from outside of the United States of America are responsible for their government import taxes and customs paperwork. Encapsula NanoSciences is NOT responsible for importation fees to countries outside of the United States of America.
- We strongly encourage the clients in Japan, Korea, Taiwan and China to order via a distributor. Tough customs clearance regulations in these countries will cause delay in custom clearance of these perishable formulations if ordered directly through us. Distributors can easily clear the packages from customs. To see the list of the distributors click here.
- Clients ordering from universities and research institutes in Australia should keep in mind that the liposome formulations are made from synthetic material and the formulations do not require a “permit to import quarantine material”. Liposomes are NOT biological products.
- If you would like your institute’s FedEx or DHL account to be charged for shipping, then please provide the account number at the time of ordering.
- Encapsula NanoSciences has no control over delays due to inclement weather or customs clearance delays. You will receive a FedEx or DHL tracking number once your order is confirmed. Contact FedEx or DHL in advance and make sure that the paperwork for customs is done on time. All subsequent shipping inquiries should be directed to Federal Express or DHL.
Storage and Shelf Life
Storage
m-Clodrosome® and m-Fluoroliposome® should always be stored at in the dark at 4°C, except when brought to room temperature for brief periods prior to animal dosing. DO NOT FREEZE. If the suspension is frozen, clodronate can be released from the liposomes thus limiting its effectiveness in depleting macrophages. ENS is not responsible for results generated by frozen product.
Shelf Life
m-Clodrosome® and Fluoroliposome® are made on daily basis. The batch that is shipped is manufactured on the same day. It is advised to use the products within 60 days of the manufacturing date.











