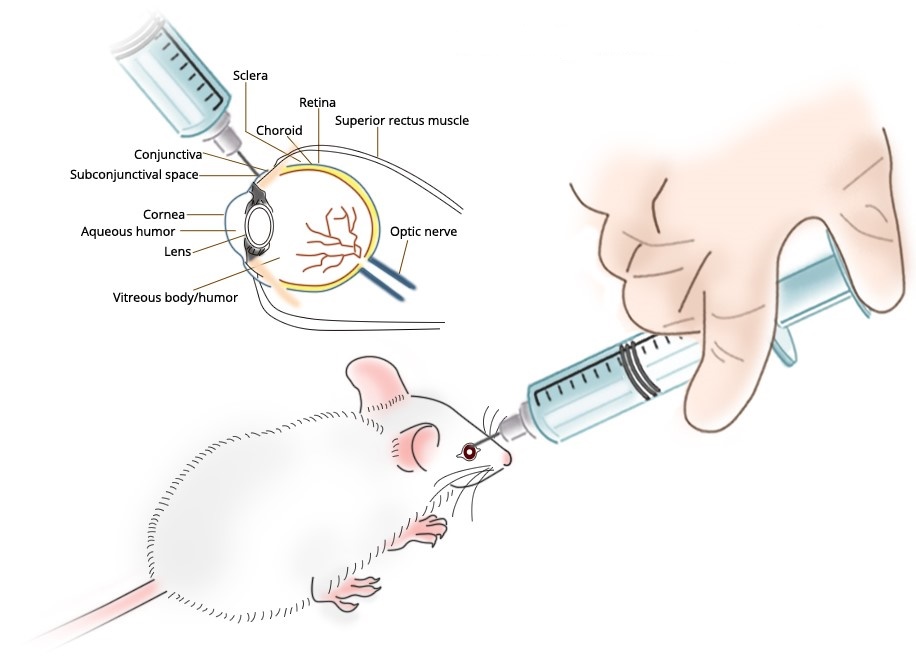
Subconjunctival Administration
Cheng et al. [1] investigated the role macrophages play in controlling herpes simplex virus (HSV)-1 replication after infection of the murine cornea. Macrophage depletion in selected tissues before or after virus infection was achieved by repeated subconjunctival (SCJ) and/or intravenous (IV) injection of liposomes containing dichloromethylene diphosphonate (L-Cl2MDP). Controls received liposomes containing phosphate-buffered saline (L-PBS). The efficiency of depletion was evaluated by histologic examination. Virus content in infected tissues was determined by standard plaque assay. Delayed-type hypersensitivity (DTH) responsiveness was assessed using the ear-swelling assay. Antibody isotype responses to virus antigens and cytokine production were monitored by enzyme-linked immunosorbent assay.
Balb/c mice given SCJ injection of L-Cl2MDP 4 and 2 days before HSV-1 corneal infection were found to have ocular virus titers as much as 105-fold higher than that seen in the L-PBS-treated controls 8 days after infection. When L-Cl2MDP treatment was delayed until 2 and 4 days after infection, virus titers in the eye were analogous to those in the control animals. Subconjunctival and submandibular lymph node macrophages in mice given local (SCJ) L-Cl2MDP pretreatment were profoundly reduced, whereas the number of corneal Langerhans’ cells and lymph node dendritic cells remained unchanged. Local L-Cl2MDP pretreatment was associated with significantly reduced DTH responsiveness to HSV-1 antigen, and an alteration in selected antibody isotype production. Depletion of macrophages in the subconjunctival tissue before corneal infection was not accompanied by enhanced virus growth at early times (2 or 4 days) after infection.
Macrophages play an important role in restricting HSV-1 growth after corneal infection. These cells appear to be required for the development of an acquired immune response, presumably by functioning in antigen processing and presentation. The hypothesis that macrophages are major participants in innate immunity to HSV-1 corneal infection was not supported.
Technical Notes
- Clodronate or lipid concentrations are stated in the paper nor does the referenced paper clearly state concentrations.
- The liposome preparation is reported by van Rooijen et al. [2].
- Subconjunctival administration of liposomal clodronate resulted in depletion of subconjunctival and submandibular macrophages (presumably due to drainage of ocular lymph into submandibular lymph nodes) but not dendritic nor Langerhans cells as judged by histological assessment of acid phosphatase+ cells.
- Subconjunctival administration of liposomal clodronate pre-infection reduced the acquired immune response (DTH) but had no effect when dosed either post-infection or intravenously.
- Subconjunctival administration of liposomal clodronate pre-infection increased the titers of anti-HSV IgG2a (2.8X) and IgM (43X).
- Clodronate liposome treatment by either route before or after infection did not affect viral production.
- Neither liver nor splenic macrophage depletion was assessed when clodronate liposomes were administered intravenously, but subconjuntival macrophages were not depleted post-i.v. treatment with clodronate liposomes.











